
Avant-propos: Alors que je rédigeais le présent message sur les crocodiliens je suis tombé sur un article du blog de M. Colin qui traitait de thématiques similaires à l'article que j'étais alors en train de rédigé. Aussi histoire de ne pas répéter ce qui a déjà été très bien expliqué par Colin, je cite certains de son article tout en mettant bien évidemment ce dernier en lien. Lisez bien son article car il y a des détails sur le métabolisme des crocodiliens qui ne sont pas mentionnés ici.
Edit: Suite à un échange sur le forum de Rationalisme.org j'ai amené quelques corrections précisions à mon présent article sur les crocodiliens. Le gros des ajouts concerne la position et le développement de l'arc aortique gauche des crocodiliens. Merci à la personne qui a souligné les erreurs/omissions de mon présent message et m'a donc permis de combler ces dernières.
_________________________________
Le titre du présent article peut sembler quelque peu fantaisiste mais avant d’expliquer de quoi il s’agit petit rappelle sur la phylogénie des crocodiliens.
Les crocodiliens sont des Archosauriens, les Archosauriens étaient jadis représentés par divers taxons, tels que les Ptérosaures et les multiples lignées de Dinosaures aujourd’hui disparues (voir l'arbre phylogénétique ci-dessous) et bien plus encore. Aujourd’hui subsistent deux importants taxons d’Archosauriens, à savoir les Dinosaures Aviens (Oiseaux) et les crocodiliens.

Phylogénie des amniotes.
Les crocodiliens d’aujourd’hui sont représentés par des taxons tels que les alligators, les gavials et bien évidemment les crocodiles à proprement parler.

De gauche à droite, un gavial, un alligator et un crocodile.
Les crocodiles sont donc des Archosauriens et sont donc plus proches des oiseaux que des autres «reptiles». Et petite précision par «autres reptiles» nous entendons bien évidemment les Lépidosauriens + les Testudines (c'est-à-dire les Tortues dont la position phylogénétique exacte parmi les amniotes demeure incertaine). D'ailleurs si l'on observe la phylogénie des amniotes ci-dessus on s'aperçoit que le terme «reptile» n'a donc pas de réelle pertinence car au mieux paraphylétique (voir aussi cet extrait du documentaire «Espèce d'Espèce»)! Certes ça beaucoup d’entre vous le savaient probablement déjà mais gardez bien cela en tête car cela est à mettre en lien avec ce qui va suivre.
Cœur de crocodile
Bon attaquons tout de suite la thématique figurant dans le titre du présent article à savoir le cœur des Crocodiliens. Pour se faire comparons schématiquement le cœur d’un Crocodilien avec celui d’un oiseau (ou d’un mammifère) ainsi qu’avec celui d’un «reptile» ou mieux dit celui d'un Lépidosaurien et/ou d'une Tortue (voir schéma ci-dessous).


Ouf vous avez tout suivit? Si c’est le cas alors jusque-là tout va bien mais attendez ce n’est pas tout puisque grâce notamment à la valve dentée et au Foramen de Panizza mentionnées ci-dessus, les crocodiliens sont en fait capables de changer leur mode de circulation sanguine! Hein quoi? Qu’est-ce que cela veut dire? L’extrait de l'article du Blog de M. Colin suivit d’une image, devrait éclairer tout le monde sur la question.
En temps normal la base de l’aorte gauche est fermée et celle-ci reçoit du sang oxygéné en provenance du ventricule gauche via le foramen de Panizza. Dans cette situation l’apport en dioxygène au reste de l’organisme est maximal.Mais lorsque l’animal est en plongée (et lorsqu’il digère), le sang désoxygéné du ventricule droit passe dans les aortes droites et gauche (via le foramen). Les poumons ne servant à rien sous l’eau leur irrigation sanguine diminue donc de façon automatique par constriction de l’artère pulmonaire (au niveau de la valve dentée). En première approximation l'ouverture de l'aorte gauche permet simplement de diminuer la résistance à l’écoulement du sang. M. Collin

Ouf voilà qui fout un coup n’est-il pas? Comme M. Colin nous venons de prendre conscience que notre cœur est bien moins performant et complexe que celui du crocodile! Mais peut-être pensez-vous que les révélations étonnantes s’arrêtent-là? Que nenni! Il nous reste à déterminer comment les crocodiliens ont pu acquérir un cœur aussi épatant!
Cœur de crocodile versus…..cœur de «reptile»!
En fin 2004 une équipe de chercheurs (Roger S. Seymour et al (2004) [1] publièrent une étude susceptible d’apporter des éclaircissements quant aux modalités d’évolution des caractéristiques si particulières du cœur des crocodiliens. Leur étude se basait sur diverses observations du développement ontogénique du cœur des crocodiliens ainsi que de celui d’autres «reptiles» c'est-à-dire de Lépidosauriens et de Tortues. Le but de pareilles observations étant notamment de déterminer à quels stades sur développement apparaissent les particularités crocodiliennes.
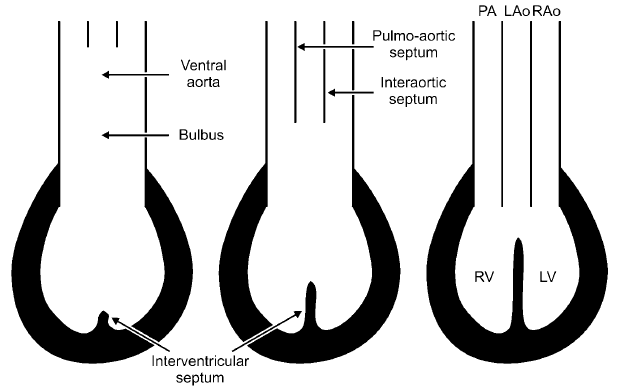

Ainsi contrairement aux autres «reptile» mais à l’instar des oiseaux le cœur des crocodiliens possède quatre cavités avec une séparation totale des cavités gauches de celles de droites. Ces observations tendant à indiquer que les premiers Archosauriens, parmi lesquels les ancêtres communs des oiseaux et des crocodiliens, avaient eux aussi un cœur dont les parties droite et gauche étaient entièrement séparées l’une de l’autre empêchant ainsi les sangs oxygéné et non-oxygéné de se mélanger dans leur cœur comme cela est pourtant le cas chez les autres «reptiles». Et cela confirmant par ailleurs le caractère désuet car non-monophylétique de la catégorie «reptiles». Cependant comme nous l’avons vu plus haut chez les crocodiliens les sangs oxygéné et désoxygéné peuvent se mélanger grâce notamment au Foramen de Panizza (celui-ci reliant l’aorte gauche à l’aorte droite) ainsi qu’à la valve dentée. La question qui se pose est de savoir comment les crocodiliens ont put acquérir un cœur aussi élaboré au cours de leur évolution.
L'étrange position de l'aorte gauche
L'aorte gauche des crocodiliens est également une curiosité puisqu'elle est relié non pas au ventricule gauche (d'où part le sang oxygéné) mais au ventricule droit (d'où part le sang désoxygéné). Ce positionnement de l'arc aortique gauche étant indispensable pour permettre aux sangs oxygéné et désoxygéné de se mélanger chez les crocodiliens. Or si l'on observe le schéma précédent on remarque que si l'arc aortique gauche part du ventricule droit cela dépend de la position du septum membraneux par apport aux trois arc aortique (arcs aortique gauche et droites ainsi que l'arc aortique pulmonaire). Seymour et al continuant leurs explications en précisant que la position des trois arcs aortiques à la sortie du cœur (qui définira en bonne parti le «mode» de circulation) dépend donc du développement ontogénique du septum membranaire!
En effet comme le souligne Seymour et al durant son développement chez les crocodiliens, le septum membranaire se développe et «s'attache» entre l'arc aortique gauche et l'arc aortique droite. Avec la conséquence que l'on connait à savoir une aorte gauche partant du ventricule droit.
Durant son développement chez les oiseaux le septum membraneux se développe et «s'attache» entre l'arc aortique gauche et l'arc aortique pulmonaire. Donc chez les oiseaux l'aorte gauche devrait prendre racine dans le ventricule gauche (Seymour et al notent toutefois quelques anomalies ponctuelles où l'arc gauche se serait malgré tout «attachée» au ventricule droit chez certains embryons d'oiseaux) . Mais attendez! Durant la suite de leur développement l'arc aortique gauche des oiseaux s'atrophie rapidement pour ne laisser place qu'au seul arc aortique droit! Et donc notons à titre de comparaison et contrairement à ce que pourrait laisser supposer ce précédent schéma, que la situation diffère de manière importante chez les mammifères, puisque chez ces derniers l'aorte qui subsiste est l'aorte gauche, là où les oiseaux retiennent donc l'aorte droite! Cela étant parfaitement logique sachant que les oiseaux sont des archosauriens tout comme les crocodiliens avec lesquels ils partagent donc l'aorte droite!
Mais quel est l'intérêt de cette comparaison crocodiliens/oiseaux? Simple sachant que durant leur développement les embryons d'oiseaux présentent un arc aortique droit «prenant racine» dans le ventricule gauche Seymour et al notent que nous pouvons assez aisément nous représenter l'évolution du cœur des crocodiliens et de celui des oiseaux à partir de celui ancêtre commun. Seymour et al ajoutant que la position de l'arc aortique gauche prenant racine dans le ventricule droit chez les crocodiliens pouvant s'expliquer par un simple «déplacement» («shift» en anglais) du septum membranaire. Pareil déplacement pouvant se faire en une fois (c'est-à-dire en une génération) Seymour et al rappelant que pareil «déplacement» du septum membraneux se produit parfois y compris chez les mammifères avec cependant-là des conséquences généralement fâcheuses pour l'animal.
L'aorte gauche des crocodiliens est également une curiosité puisqu'elle est relié non pas au ventricule gauche (d'où part le sang oxygéné) mais au ventricule droit (d'où part le sang désoxygéné). Ce positionnement de l'arc aortique gauche étant indispensable pour permettre aux sangs oxygéné et désoxygéné de se mélanger chez les crocodiliens. Or si l'on observe le schéma précédent on remarque que si l'arc aortique gauche part du ventricule droit cela dépend de la position du septum membraneux par apport aux trois arc aortique (arcs aortique gauche et droites ainsi que l'arc aortique pulmonaire). Seymour et al continuant leurs explications en précisant que la position des trois arcs aortiques à la sortie du cœur (qui définira en bonne parti le «mode» de circulation) dépend donc du développement ontogénique du septum membranaire!
En effet comme le souligne Seymour et al durant son développement chez les crocodiliens, le septum membranaire se développe et «s'attache» entre l'arc aortique gauche et l'arc aortique droite. Avec la conséquence que l'on connait à savoir une aorte gauche partant du ventricule droit.
Durant son développement chez les oiseaux le septum membraneux se développe et «s'attache» entre l'arc aortique gauche et l'arc aortique pulmonaire. Donc chez les oiseaux l'aorte gauche devrait prendre racine dans le ventricule gauche (Seymour et al notent toutefois quelques anomalies ponctuelles où l'arc gauche se serait malgré tout «attachée» au ventricule droit chez certains embryons d'oiseaux) . Mais attendez! Durant la suite de leur développement l'arc aortique gauche des oiseaux s'atrophie rapidement pour ne laisser place qu'au seul arc aortique droit! Et donc notons à titre de comparaison et contrairement à ce que pourrait laisser supposer ce précédent schéma, que la situation diffère de manière importante chez les mammifères, puisque chez ces derniers l'aorte qui subsiste est l'aorte gauche, là où les oiseaux retiennent donc l'aorte droite! Cela étant parfaitement logique sachant que les oiseaux sont des archosauriens tout comme les crocodiliens avec lesquels ils partagent donc l'aorte droite!
Mais quel est l'intérêt de cette comparaison crocodiliens/oiseaux? Simple sachant que durant leur développement les embryons d'oiseaux présentent un arc aortique droit «prenant racine» dans le ventricule gauche Seymour et al notent que nous pouvons assez aisément nous représenter l'évolution du cœur des crocodiliens et de celui des oiseaux à partir de celui ancêtre commun. Seymour et al ajoutant que la position de l'arc aortique gauche prenant racine dans le ventricule droit chez les crocodiliens pouvant s'expliquer par un simple «déplacement» («shift» en anglais) du septum membranaire. Pareil déplacement pouvant se faire en une fois (c'est-à-dire en une génération) Seymour et al rappelant que pareil «déplacement» du septum membraneux se produit parfois y compris chez les mammifères avec cependant-là des conséquences généralement fâcheuses pour l'animal.
Le Foramen de Panizza est-il plésiomorphe ou apomorphe?
La question qui se pose ensuite est «l’origine évolutive» d'une autre caractéristique propre au cœur des crocodiliens à savoir le Foramen de Panizza. Ainsi par exemple dans son blog Jean-Philippe Colin souligne une théorie voulant que le Foramen de Panizza soit plésiomorphe c’est-à-dire un caractère ancestral présent chez les premiers Archosauriens.
Pour beaucoup, cette petite communication [le Foramen de Panizza] est un caractère ancestral conservé (plésiomorphe) qui n’est pas adaptatif, en gros « c’est comme ça, c’était déjà là, et ça sert à rien… ». Les seuls organismes dont le cœur est parfaitement cloisonné étant des animaux à sang chaud ou "homéothermes" (mammifères et oiseaux). M. Colin
L’hypothèse selon laquelle le Foramen de Panizza serait un caractère plésiomorphe est tout à fait crédible et signifierait que les premiers Archosauriens (parmi lesquels les ancêtres communs des crocodiles et des oiseaux) voyaient probablement déjà leurs sangs oxygéné et désoxygéné se mélanger. Mais c’est justement-là un point que «conteste» l’étude de Seymour et al en se basant sur les modalités de développement du cœur des crocodiliens. En effet certains se souviendront peut-être ici en quoi la biologie du développement nous renseigne sur l’évolution des animaux, au point que l’on parle souvent d’évo-dévo. Sans réexpliquer ici toutes les subtilités qui entourent la notion d’évo-dévo, rappelons simplement que généralement (mais pas toujours) les caractéristiques anatomiques qui apparaissent tôt durant le développement d’un organisme sont celles qui sont les plus anciennes (donc que possédaient déjà les lointains ancêtres de l’organisme) à l’inverse un caractère qui apparait tardivement durant le développement est davantage susceptible d’être un caractère apparu récemment dans l’histoire évolutive de l’organisme en question. Mais attention de ne pas tomber dans le piège d’un récapitulationnisme naïf voulant que l’ontogenèse récapitule parfaitement la phylogenèse. Souvenons-nous du modèle de développement en sablier [2] ainsi que des cas de Néoténie. Ces précisions en étant faites voyons ce que nous avons à apprendre sur le développement et l’évolution du Foramen de Panizza.
Dans leur étude Seymour et al relatent tout d’abord les thèses de certains chercheurs tels que Goodrich (1958) qui pensaient que le Foramen de Panizza apparaissait simplement parce que durant le développement du cœur des crocodiliens, le septum inter-aortique séparent l’aorte gauche de l’aorte droite, se développerait de manière incomplète laissant donc une «ouverture» entre les deux arcs aortiques, ouverture qui serait donc le Foramen de Panizza. Cette théorie est compatible avec l’idée que le Foramen de Panizza est un caractère «primitif» c’est-à-dire plésiomorphe. Cependant Seymour et al soulignent que même si Goodrich avait raison cela ne s’opposerait non-plus pas à l’idée selon laquelle le Foramen de Panizza serait un caractère dérivé, le développement incomplet du septum inter-aortique pouvant être perçu comme un caractère néoténique secondairement acquis au cours de l’évolution des crocodiliens.
Mais finalement Roger S. Seymour et al relatent des observations qui ne collent pas avec la théorie de Goodrich. En effet les observations en question soulignent que le Foramen de Panizza apparait après que le septum inter-aortique se soit formé. Cela tend à appuyer l’idée selon laquelle le Foramen de Panizza serait un caractère «dérivé», c’est-à-dire une innovation évolutive propre aux crocodiliens et non pas un caractère «primitif» déjà présent chez les premiers Archosauriens et dont auraient hérité les crocodiliens actuels.

Et la valve dentée?
Oui la valve dentée, un peu plus et on l’oubliait cette petit coquine! Rappelons-nous que la valve dentée est l'une des caractéristiques du cœur des crocodiliens qui permet de «switcher» leur circulation sanguine.
Que nous apprennent Seymour et al concernant la valve dentée? Concernant celle-ci les observations sont même des plus révélatrices que celles concernant le Foramen de Panizza puisque durant tout le développement et même à l’éclosion les crocodiliens analysés n’avaient pas de valves dentées pleinement formées et fonctionnelles. En réalité la valve dentée termine de se développer après la naissance de l’animal et elle ne sera pleinement fonctionnelle qu’une fois son développement terminé. Cela indiquant que la valve dentée est probablement elle aussi un caractère dérivé acquis tardivement durant l’évolution des crocodiliens!
Bien évidemment une personne avisée en biologie de l’évolution peut rappeler la nécessaire prudence à avoir face à ces observations en rappelant que l’ontogenèse ne récapitule pas (parfaitement) la phylogenèse. Néanmoins ces observations constituent indéniablement des éléments en faveur d’une origine «tardive» du Foramen de Panizza et de la valve dentée. Cela renforçant une autre hypothèse à savoir celle voulant que les premiers Archosauriens, parmi lesquels les ancêtres communs des crocodiliens et des oiseaux, possédaient déjà un cœur cloisonné entre sa partie gauche et sa partie droite et que donc les sangs oxygéné et non-oxygéné ne se mélangeaient pas. Ainsi la capacité des crocodiliens de «switcher» leur circulation sanguine donc de mélanger leurs sangs oxygéné et non-oxygéné serait un caractère dérivé c’est-à-dire une apomorphie!
Seymour et al soulignant qu’il est alors fort probable que les premiers Archosauriens étaient des animaux où pareil mélange des sangs oxygéné et non-oxygéné n’avaient pas lieu et que donc les premiers Archosauriens auraient été homéothermes. Cela impliquerait alors que le métabolisme ectotherme des crocodiliens actuels représenterait lui aussi un caractère «dérivé» acquis par les crocodiliens au cours de leur évolution.
Or tout cela implique que les ancêtres des crocodiliens étaient peut-être à sang chaud! Pourquoi? Simplement parce qu’il est donc fort probable que les ancêtres des crocodiliens avaient déjà un cœur dont les cavité gauches sont totalement séparées de celles de droites, mais n'avaient donc pas ni de Foramen de Panizza, ni d'aorte gauche partant du ventriculaire droit, ni valve dentée. Cela impliquant que les dits ancêtres ne pouvaient donc probablement pas mélanger le sang oxygéné au sang désoxygéné. Or ce non-mélange est généralement associé à l’homéothermie. [3] Roger S. Seymour et al soulignent que ce scénario a rarement été considéré dans le passé en raison d’une certaine préconception voulant que les homéothermes, vues à tort comme étant plus « évolués», dérivent des ectothermes et non l’inverse. Mais donc contrairement à cette préconception Roger S. Seymour et al rappelle que des cas d’évolution dans «l’autre sens» ont dû exister et existent, les crocodiliens n’en seraient donc qu’un exemple parmi d’autres.
Et ce n’est pas tout!
Notons que le cœur exceptionnel des crocodiliens ainsi que les modalités de développement de celui-ci ne sont pas les seuls indices en faveur d’ancêtres Archosauriens homéothermes. Par exemple saviez-vous que le système respiratoire des crocodiliens est étrangement similaire à celui des oiseaux? [4]
Mieux encore le registre fossile nous amène de précieuses informations sur ce à quoi pouvaient ressembler les ancêtres des crocodiliens. En effet rappelons-nous que les crocodiliens ne sont qu’un rameau d’un taxon d’Archosauriens jadis bien plus diversifié et que l’on nomme les Crocodylomorpha. En effet les fossiles de ces anciens cousins de nos crocodiliens actuels tendent à montrer qu’ils avaient pour beaucoup un mode de vie terrestre et contrairement aux crocodiles actuels des membres bien redressés sous leur corps voir même pour certains, adoptaient une locomotion bipède [5]. Le positionnement de leurs membres et leur locomotion étaient en effet plus similaire à celui des dinosaures qu’à celui de nos crocodiliens actuels, tout cela étant compatible avec un mode de vie actif lui-même compatible avec l’homéothermie!

Conclusion:
Même si il n’y a à ce jour aucune certitude, la prise en compte des liens de parentés réels des crocodiliens ainsi qu’une meilleure connaissance de leur biologie, nous oblige à considérer avec sérieux la théorie selon laquelle les ancêtres des crocodiliens actuels aient été homéothermes.
Ces nouvelles perspectives et théories sur l’évolution des crocodiliens en particulier et des Archosauriens en général, sont également là pour nous rappeler que l’homme et les autres mammifères avec leur homéothermie et leur cœur à quatre cavités ne sont pas le «summum» en matière d’évolution. Les crocodiliens ont un cœur bien plus «complexe» et «élaboré» que le nôtre et nous rappellent non seulement que nous ne sommes pas au sommet de l’échelle de l’évolution mais également que l’échelle de l’évolution n’existe tout simplement pas.
Références:
[1] Roger S. Seymour et al (2004), Evidence for Endothermic Ancestors of Crocodiles at the Stem of Archosaur Evolution, The University Chicago Press
[2] Naoki Irie & Shigeru Kuratani (2011), Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis, Nature
[3] Adam P. Summers (2005), Evolution: Warm-hearted crocs, Nature
[4] K. T. Bates and E. R. Schachner (2011), Disparity and convergence in bipedal archosaur locomotion, Journal of The Royal Society
[5] C. G. Farmer and Kent Sanders (2010), Unidirectional Airflow in the Lungs of Alligators, Science
J'ai un ami qui fait une thèse sur les crocodiles (actuels et fossiles) pour savoir s'ils avaient primitivement effectivement le sang chaud ou non... Suspens ! Même si tout ça y laisse penser on ne sait jamais, ça pourrait-être des exaptations...
RépondreSupprimerOui il n'y aura probablement jamais de certitudes ce qui est pour le moins rageant quant on y pense. Soit dit en passant Mea culpa si je me montre pour le moins partial en faveur d'ancêtres homéothermes pour les crocodiliens.
RépondreSupprimer