Alors bon je me suis finalement décidé à faire une petite mise au point à l’attention de Meng Hu, sur l’idiotie de ses propos (et de sa personne) en revenant justement sur cet exemple spécifique du chromosome Y. Et pourtant Dieu sait que ce n'est de loin pas la seule bouffonnerie susceptible d'être mise en avant dans ce pavé de conneries retranscrit par le dénommé Meng Hu.
____________________________________________________________________
Salut Meng Hu ça va?
Bon je ne vais pas m’attarder davantage en insipides formules de politesse pour attaquer directement le beefsteak si tu veux bien.
Tu es un raciste qui s’est amusé à retranscrire de manière détaillée la diarrhée thèse d’un certain Richard D. Fuerle. J’ai déjà lu de nombreux étrons écrits racialistes, mais alors ceux de Fuerle c’est le pompon! Il faut dire Meng Hu que lorsque j’avais lu (en diagonale je l’avoue mais cela m’a amplement suffit) ta retranscription de «l’œuvre» de Fuerle sur ton blog, je m’étais bien marré. Ce qui m’a fait marrer n’était pas tant les habituels excréments racistes que l’on connait (notamment ceux consistant à rapprocher les noirs des singes) mais surtout le «scénario évolutif» neuneu nommé «Out of Eurasia».
En effet un des éléments centraux de la chiasse thèse de Fuerle étant que les africains n’aient jamais quitté leur continent autrement qu’en tant qu’esclaves déportés de force en Eurasie puis plus tard en Amérique. Fuerle rejetant totalement l’idée que l’homme moderne ait pu apparaitre en Afrique pour ensuite quitter cette dernière et coloniser l’Eurasie et le reste du monde (cela allant de pair avec le racisme viscéral du bonhomme). Il faut dire mon cher Meng Hu qu’après avoir lu ta retranscription de la crotte thèse de Fuerle je n’ai bien évidemment pas penser à répondre à la dite «thèse» tant elle atteint des sommets niveau connerie et tant j’ai mieux à foutre de mon précieux temps que de consacrer celui-ci à tous les pavés de conneries pondu par des zozos racistes qui débitent tout n’importe quoi sur le Net…..
Néanmoins alors que j’étais en train de rédiger le prochain message de mon blog consacré aux haplogroupes du chromosome Y je me suis souvenu d’un point particulier de ta retranscription de la bouse thèse de Fuerle.
Mais avant tout mon cher Meng Hu je me permet de citer
un échange que tu avais eu avec une personne trouvant à juste titre la
merde thèse de Fuerle ridicule à souhait.
babar John: “Ceux sur la “théorie” de fuerle qu’il a nommée Out of Eurasia sont de loin les plus ridicules selon moi.”
Meng Hu: Je vous souhaite bonne chance pour lancer un commentaire sur la centaine de références citées. Et n’allez surtout pas croire que s’attaquer à la Section 4 revient à réfuter la Section 3 : celle qui réfute directement Out of Africa.
Ce commentaire de ta part m’a beaucoup amusé Meng Hu car il me rappelle ceux d’un créationniste qui me disait un peu-près la même chose avec
l’Atlas de la Création de
Harun Yahya. Tu penses bien que s’amuser à réfuter point par point de pareils pavés de conneries n’est pas à la portée de tous, car hormis avoir du temps libre à ne plus savoir qu’en foutre il faut aussi être sacrément maso pour perdre son temps à lire et à réfuter dans les détails les pavés délirants issus d’on ne sait quel taré.
Néanmoins alors que je rédigeais
mon billet sur les haplogroupes du chromosome Y, je me suis décidé de revenir sur un point particulièrement amusant de la soit disant «réfutation» de «Out of Africa» par Richard D. Fuerle. Et c’est aussi amusant par apport à toi mon cher Meng Hu, car cela démontre également ton ignorance crasse en matière de génétique (et plus généralement en matière de paléoanthropologie) ainsi que ta capacité à gober n’importe quelle connerie à partir du moment qu’elle va dans le sens de tes préjugés raciaux à deux roupies. Mais bon aller trêve d’avant-propos et allons dans le vif du sujet en revenant
sur ta retranscription des propos de Richard D. Fuerle concernant le chromosome Y.
Parallèlement, l’ADN du chromosome Y est transmis par la lignée masculine et nous montre où les hommes voyageaient. Il ne raconte pas exactement la même histoire, étant donné que les hommes ont beaucoup plus exploré que les femmes. Les hommes se déplaçaient souvent vers de nouvelles terres sans leurs femmes, ensuite, ils s’accouplèrent avec des femmes indigènes, de sorte que leurs descendants avaient l’ADNmt des natifs et l’ADN du chromosome Y des explorateurs.
La carte ci-dessus montre la répartition mondiale des différentes variations du chromosome Y. La quantité de chaque couleur dans les cercles est proportionnelle au nombre d’hommes dans cet endroit qui avait la variation indiquée par cette couleur. On note que l’olive, la couleur majeure en Afrique, apparaît à l’extérieur de l’Afrique, mais seulement autour de la Méditerranée, ce qui suggère que les africains n’ont pas migré hors d’Afrique, sauf en tant qu’esclaves emmenés dans ces zones. Le rouge vif et le bleu foncé sont uniques à l’Afrique, ce qui suggère également qu’il n’y avait pas de migrations hors d’Afrique. Ces variations peuvent avoir été apportées en Afrique par les hominoïdes primitifs (section IV), qui se sont éteints ailleurs, mais dont l’ADN-Y persiste encore en Afrique.
Les couleurs européennes orange et jaune indiquent que des hommes européens vivaient dans le Moyen-Orient, l’Afrique du Nord, la Géorgie de l’ex-URSS, l’Inde, l’Asie du Sud-Est, l’Australie et l’Amérique du Nord (les hommes en orange et jaune peuvent avoir été les membres d’une seule population). Le vert est la couleur dominante dans les Amériques, et les petites quantités de vert dans l’Ancien Monde indiquent leurs origines en Asie de l’Ouest, migrant par la suite vers le nord de l’Inde et le sud de la Sibérie et, éventuellement, les Aïnous au Japon.
Du fait des grandes quantités de rose dans l’est de l’Asie, on pourrait s’attendre à d’importantes quantités de rose dans les Amériques, mais elle n’est pas là, ce qui suggère que les asiatiques en rose ne sont pas enclins à beaucoup explorer et que moins d’évolution s’est produite chez les asiatiques en rose que chez les européens en orange et jaune. Meng Hu

Dis-moi Meng Hu, à ton avis sur une échelle de 1 à 10 quel est le degré de débilité de ta présente prose sur les variations du chromosome Y? Personnellement je dirais 100 ou 101 et encore je suis gentil!
Il faut dire que seul un ignare aurait pu lire de manière aussi
stupide naïve la carte de répartition des haplogroupes du chromosome Y. Et toute personne ayant été quelque peu familiarisée en ce qui concerne les haplogroupes en question, sait que la présente carte ci-dessus est insuffisante pour comprendre comme il se doit la question des haplogroupes des différentes populations humaines. Mieux lorsque l'on se réfère à
l'étude originale de P.A. Underhill et al (2001) [1] dont est issue cette carte on s'aperçoit que le dit Richard D. Fuerle l'a sorti de son contexte en omettant le diagramme qui va avec (voir image ci-dessous)!

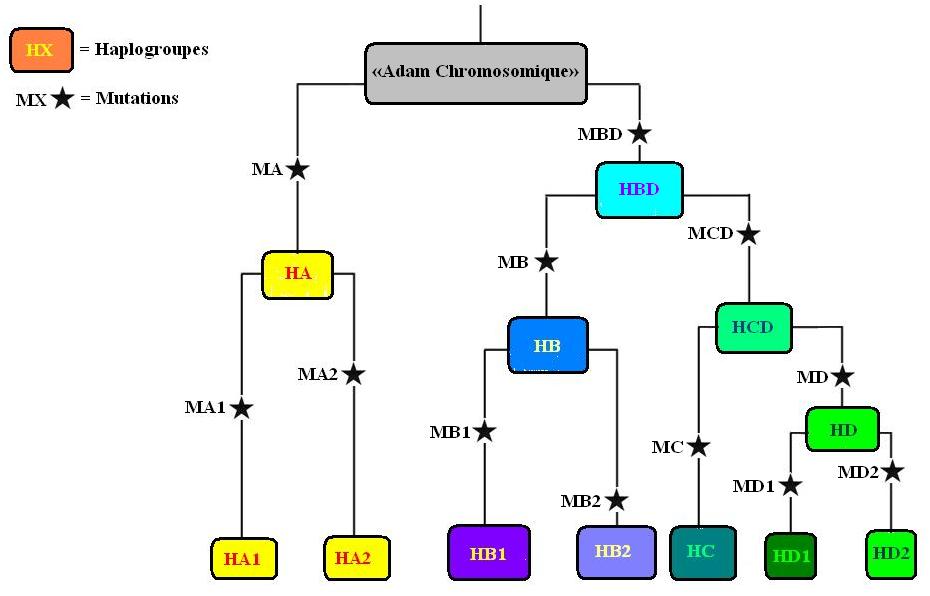
Ci-dessus la carte issus de l’étude de Underhill et al (2001) [1], accompagné comme il se doit du diagramme qu’a (volontairement?) omit Richard D. Fuerle. Ce diagramme est un arbre d’apparentement (phylogénétique) des différents haplogroupes du chromosome Y. Si l’on prend l’ensemble des embranchements représentés en couleur olive, on remarque que ces derniers appartiennent à un plus grand embranchement auquel appartient également les haplogroupes de couleur bleu clair. Mais ce dernier embranchement appartient également à un embranchement encore plus vaste réunissant également tous les embranchement de droite (allant de V à X). Mais ce grand embranchement appartient à un embranchement encore plus grand réunissant également les embranchements représentés en bleu foncé. Ce plus grand embranchement étant lui-même un sous-ensemble d’un embranchement encore plus vaste comprenant également les embranchements en rouge tout à gauche du diagramme.
Une fois la carte de répartition des haplogroupes remise dans son contexte on remarque que les divers embranchements sont des sous-ensembles d’embranchements plus vastes. Nous obtenons donc un véritable
dendrogramme dont les embranchements les plus basaux ainsi que la localisation de ces derniers, peuvent nous permettre d’estimer l’origine géographique probable du dernier ancêtre patrilinéaire commun aussi parfois appelé
«Adam chromosomique». Le but étant donc de savoir où s’enracine l’arbre en question.
Mais vu que tu ne comprends probablement toujours pas le Schmilblick mon très cher Meng Hu, je te conseille de te référer à
ce message où j’explique plus en détail la manière dont on définit les haplogroupes du chromosome Y et quels enseignements peuvent-être tirés de ces derniers.
Et c’est d’ailleurs en lisant
le message en question que tu t’apercevras que la carte cité par Richard D. Fuerle, montre exactement le contraire de ce que celui-ci prétend! Car les haplogroupes en rouges et en bleu sont les plus basaux de l’arbre d’apparentement des haplogroupes. Cela signifie que l’arbre s’enracine en Afrique et que l’ensemble des haplogroupes non-africains témoignent en réalité de migrations en provenance du continent africain. De plus nous parlons de sorties relativement récentes. Par exemple l’embranchement contenant l’ensemble des haplogroupes à l’exception des haplogroupes rouges tout à gauche, représente à lui seule
l’haplogroupe BT. Si l’on se réfère à une estimation ancienne attribuée à cet haplogroupe, nous avons une date de 75'000 ans
Cruciani et al (2011) [2]. Et donc cet haplogroupe BT s’enracinant clairement sur le continent africain, celui-ci témoigne donc de sorties d’Afrique largement plus récentes qu’il y a 75'000 ans!

Distribution hiérarchique des haplogroupes. Les embranchements (haplogroupes) A, A1a-T, A2-T, et BT, s’enracinent clairement sur le continent Africain. Certains pensent que l’haplogroupe DE aurait pu apparaitre en Eurasie suite à une première sortie du continent africain (sortie qui aurait eu lieu il y a moins de 100'000 ans) puis se serait répandu en Afrique suite à un retour sur le continent africain de certaines de ces populations humaines. Selon ce dernier scénario l’haplogroupe E, très commun en Afrique, serait donc le fruit d’importants «retours» sur le continent africain de populations humaines dont les ancêtres avaient récemment quitté ce dernier. Ce scénario est tout à fait plausible. Cependant le scénario le plus parcimonieux et que l’haplogroupe DE soit apparu en Afrique (voir mon message consacré aux haplogroupes du chromosome Y).
Note mon Meng Hu, l’haplogroupe E c’est l’haplogroupe couleur olive représenté sur la carte de répartition géographique des haplogroupes que tu avais commenté. Et c’est via cette carte qui montre la répartition de l’haplogroupe E, que Richard D. Fuerle (et toi-même) affirme que les africains n’auraient jamais quitté leur continent autrement qu’en tant qu’esclaves importés de force en Eurasie.
La débilité de l’affirmation de Richard D. Fuerle, est donc ici maximale sachant que l’haplogroupe E, à l’instar des divers haplogroupes eurasiatiques, sont tous des sous-ensembles d’haplogroupes plus vastes (par exemple de l’haplogroupe BT largement plus récent que 100'000 ans) et s’enracinant en Afrique.
Par ailleurs le plus croustillant c’est quand dans
la section IV du «dossier» que tu as consacré aux
immondices idées de Richard D. Fuerle sur ton blog tu cites des publications dont tu ne captes manifestement que pouic! En effet dans cette section IV tu cites trois extraits d’article sensés validés une «origine eurasiatique»
des Khoisans et probablement par là-même constitué un élément de preuve à la
déjection thèse «Out of Eurasia» de Richard D. Fuerle. Seulement voilà il y a deux
bouffonneries problèmes majeurs dans cette «présentation».
Premièrement le fait que les Bushmen puissent être issus d’un mélange entre des populations humaines restées en Afrique et d’autres effectuant un retour en Afrique depuis le Proche-Orient, n’est pas un élément de preuve de la chiure théorie «Out of Eurasia» telle que présentée par Richard D. Fuerle, sachant que l’on parle simplement ici, au mieux, de populations humaines relativement récemment sortie d’Afrique qui y seraient retournés en se mélangeant avec des Homo sapiens qui eux n’ont pas quitté le continent africain.
Deuxièmement mon Meng Hu, force est de constater que tu ne comprends pas ou pire encore, ne lis pas les sources que tu cites. Démonstration avec les extraits suivants
que tu as posté…..
Phylogeographic analyses suggest that a large component of the present Khoisan gene pool is eastern African in origin and that Asia was the source of a back migration to sub-Saharan Africa [...] sub-Saharan Africa is represented by a set of chromosomes that harbors the M207 and M173 mutations [...] The last scenario, that of a back migration from Asia to Africa, currently appears to be by far the most plausible. This is because most of the M9 haplotypes (the majority of group VII and VIII lineages, as well as some group IX and X lineages reported by Underhill et al. [2000]) have been observed only in Asia. Moreover, this possibility appears to be further supported by the recent finding of the UTY2+/M173- intermediate haplotype (Karafet et al. 2001) in central and northeastern Asia (the UTY2 marker in the study by Karafet et al. [2001] corresponds to M207 in the present study). (Cruciani et al., 2002)
Tu sais Meng Hu quand on en arrive à ne capter le quart du tiers d’une publication scientifique (ici une publication de
Fulvio Cruciani et al (2002) [3]), il est préférable de s’abstenir de la citer comme démonstration de son point de vue! Dans cette citation tu as en effet coupé et mis ensemble divers extraits qui ne traitent pas directement de la même chose. En effet la première phrase que tu cites est issue de l'abstract et concerne de précédentes études émettant l’hypothèse qu’une partie des ancêtres des Khoisans auraient été des populations d’homme modernes ayant effectués une migration retour en Afrique depuis le Proche-Orient.
Mais les autres extraits (issue de la même étude) que tu as cité ci-dessus, ne concernent pas les Khoisans! En effet les extraits sur les mutations M207 (UTY2) et M173 concernent en réalité un haplogroupe que l’on nomme aujourd’hui
Haplogroupe R et plus spécifiquement
Haplogroupe R1. Et l’étude que tu cites, explique clairement que cet haplogroupe se trouve fréquemment dans une région aux alentours du Nord du Cameroun et non pas chez les Khoisans! De plus l’haplogroupe R est relativement récent (largement plus récent que 50'000 ans) et donc sa présence dans la région traduit simplement des vagues migratoires récentes largement ultérieures à l’apparition de l’homme moderne, dont les haplogroupes plus anciens nous montrent des sorties d’Afrique des populations d’homme modernes.
Mais le plus amusant est qu’après avoir cité cette source sans la comprendre tu en cites à nouveau une autre que tu présentes toujours dans le cadre de la «démonstration» d’une origine asiatique des Khoisans…..
… the possibility that YAP haplotype 3 originated in Asia and migrated to Africa. This hypothesis is supported by the finding of high frequencies of haplotype 3 in some Asian populations (i.e., ~ 50% in Tibet) and by the observation of higher levels of diversity (based on the number and frequency of alleles at the DYS19 microsatellite locus) associated with Asian versus African haplotype 3 chromosomes. Because YAP haplotypes 4 and 5 evolved from haplotype 3 and account for the majority of Y chromosomes in Africa (table 1), this hypothesis implies a substantial Asian contribution to the African paternal gene pool (Hammer et al. 1997). (Altheide & Hammer, 1997)
Là c’est sûr et certain Meng Hu, ni toi ni Richard D. Fuerle ne captez quoi que ce soit de la thématique que vous abordez! Car voilà le présent article de
Altheide & Hammer, (1997) [4] est contredite par l’étude de Fulvio Cruciani
et al (2002)
[3] que tu avais cité juste avant. En effet dans l’étude de Fulvio Cruciani
et al (2002)
[3] on peut lire ceci:
An ancient human back migration from Asia to Africa had already been proposed by Altheide and Hammer (1997) and Hammer et al. (1998, 2001), on the basis of nested cladistic analysis of Y-chromosome data. They suggested that the presence of YAP_ chromosomes in Africa was due to such an event, but this has recently been questioned by Underhill et al. (2001b) and Underhill and Roseman (2001), primarily on the basis of the Asian-specific YAP_ subclade that neutralizes the previous phylogenetic inferences. Thus, the only evidence of a migration from Asia to sub-Saharan Africa that is fully supported by Y-chromosome data relies, at least for the moment, on the finding of haplogroup IX chromosomes in Cameroon. (Cruciani et al., 2002)
Comment expliquer une telle omission de ta part et de celle de Richard D. Fuerle mon cher Meng Hu? Entre la malhonnêteté et la stupidité mon cœur balance…..
Et cela en ajoutant que la mutation YAP définit en réalité l’haplogroupe DE dont l’un des sous-haplogroupe n’est autre que l’haplogroupe E. Or tu te souviens Meng Hu, l’haplogroupe E c’est l’haplogroupe représenté en «couleur olive» dont tu affirmais que la répartition prouvait une absence de migration des africains hors d’Afrique autrement qu’en tant qu’esclaves déportés de force. Or sans le savoir tu balances une référence qui stipule que l’haplogroupe DE et de ce fait l’haplpogroupe E, serait orignaire d’Asie….. Bref Richard D. Fuerle et toi-même vous enfoncez dans des auto-contradictions et incohérences pour le moins énormes. Preuve mon cher Meng Hu que tu ne lis pas les références ou que tu ne piges strictement rien aux références auxquelles tu te réfères!
Mais surtout que l’origine géographique de la mutation YAP soit africaine ou asiatique cela ne change rien au fait que l’arbre phylogénétique du chromosome Y montre clairement une expansion humaine hors d’Afrique il y a moins de 100'000 ans! L’haplogroupe DE définit par la mutation YAP n’en demeurant pas moins un sous-ensemble d’haplogroupes plus vastes enracinés en Afrique. Tout cela étant donc en réalité et en dépit des délires de Richard D- Fuerle, une démonstration d’une expansion de l’homme moderne à travers le monde à partir du continent africain!
Et ce genre de conneries de ta part Meng Hu est d'autant plus croustillante lorsque l'on sait
que tu reproches à d'autres de ne pas lire les sources du dénommé Richard D. Fuerle.
Sérieusement, vos posts me renforcent dans l’idée que vous n’avez pas lu le livre et la centaine de références citées, ni même les sections 1 et 2 contrairement à ce que vous prétendez. Je fatigue. Diable, que vais-je faire de vous ? Meng Hu
Arf, arf, arf, et donc mon Meng Hu que dois-je déduire concernant ta petite personne? Que tu n'es même pas foutu de capter les références que tu cites? Ou alors pire que tu ne les lis pas toi-même? En tout cas merci de confirmer que tu n'es qu'un imbécile inconséquent qui reproches à d'autres une stupidité dont tu fais toi-même preuve à tour de bras!
Et donc voici un bon conseil Meng Hu, avant de jouer le petits donneurs de leçons suffisants commence par te trouver un cerveau en état de marche car le tient est de toute évidence périmé et/ou hors service. Après et seulement après tu comprendras peut-être pourquoi la sélection de publications à dessein (ici à dessein idéologique et plus exactement raciste) est une stratégie aussi idiote que malhonnête sur le plan intellectuel (c’est d’ailleurs également la stratégie favorite des créationnistes).
Il faut dire que le résultat de pareille «démarche» est toujours le même à savoir un patchwork d’affirmation fallacieuses, s’appuyant sur des extraits de publications qui sont soit boiteuses (et ce n’est pas cela qui manque en Science), ou alors soit correctes, mais non comprises par la tête de nœud qui en citent des extraits qui sorties de leur contexte, entrent dans la catégorie
«Quote Mining». Richard Fuerle et donc toi-même Meng Hu, entrant tous deux dans cette dernière.
Mais donc Meng Hu comment expliquer pareilles faussetés, auto-contradictions et démonstrations d’ignorance dans les textes de Richard D. Fuerle? Comment peux-tu toi-même faire preuve de pareille stupidité en retranscrivant ceux-ci?
La réponse à cette dernière question est évidente pour toute personne qui sait distinguer un texte scientifique d’un ramassis de conneries idéologiques! En effet qui serait assez con à part quelques gogols comme Meng Hu pour prendre comme référence scientifique un texte où l’on peut lire ceci?
Today, white men in the military fight all over the world, but they do not fight for the one thing that is most important to the survival of their kind – who impregnates their women. They not only condone the impregnation of white women by other races, they not only facilitate it, they actually celebrate it! Richard D. Fuerle
Putain j'ai vraiment failli m'étrangler de rire en lisant cette prose. On parie combien que la tension du dénommé Richard D. Fuerle monte dangereusement lorsqu’il croise une jolie blonde accompagnée d’un noir?
Mais donc je ne vais quand même pas commenter l’ensemble des inepties racistes venant de crétins décérébrés même pas foutus de capter les publications auxquelles ils se réfèrent (et encore je pars de l’idée généreuse voulant qu’ils aient lu les dites publications). Ainsi Meng Hu pour l’ensemble tes inepties racistes tels que celle-ci…
Le bien le plus précieux que les blancs possèdent, c’est leur génome. Ils peuvent perdre des territoires et des richesses, mais si leur génome est intact, ils peuvent survivre et récupérer tout ce qu’ils ont perdu. Si leur génome est souillé, il n’y a plus de retour possible. Meng Hu
...je me contenterai de te répondre par ceci...



























{kind=link}